Course Details of Training Workshop on Biology and Ecology of Copepods
 The training course will last five days and will comprise a combination of lectures, laboratory-based practical sessions and informal, problem-solving discussions and cruises with a hatcheries tour on the final day. Flexibility will be maintained to encourage the development of an intense group dynamic through interactions between highly motivated young researchers and established professionals. Participants will be requested to bring biological specimens or molecular data with them that are of special interest or significance in their own work – particularly their problem species, which will be
examined in practical sessions together with relevant tutors. Questions concerning molecular analyses will be dealt with by Dr. Iole Di Capua.
The training course will last five days and will comprise a combination of lectures, laboratory-based practical sessions and informal, problem-solving discussions and cruises with a hatcheries tour on the final day. Flexibility will be maintained to encourage the development of an intense group dynamic through interactions between highly motivated young researchers and established professionals. Participants will be requested to bring biological specimens or molecular data with them that are of special interest or significance in their own work – particularly their problem species, which will be
examined in practical sessions together with relevant tutors. Questions concerning molecular analyses will be dealt with by Dr. Iole Di Capua.
Practical training on board the university-owned survey ship TRV Toyoshio-maru (Hiroshima University) will be provided for all students. Participants will be introduced to the use of various sampling gear and experience life on board. Near FL, seagrass beds and mud flats provide the opportunity to observe tri-spine horseshoe crabs (Tachypleus tridentatus) and collect phytal and benthic copepods. The workshop will finish with an excursion to the local Hiroshima Prefectural Hatcheries Laboratory on Friday 31 May.
Download the syllabus of this training workshop by clicking the image. You can also read the syllabus below.
Lectures Training Cruise Practical Session Hatchery Tour Others
Lectures
Morning of Day 2 (27 May 2024)
Lecture 1-1: Introduction to Copepod Morphology, Ordinal Classification and Phylogeny (Tutor: Rony Huys)
 This session will provide a brief overview of copepod morphology and diversity and introduce
participants to the impressive range of body forms and limb types across the Copepoda. Trainees will be introduced to the standardized terminology used in the description and identification of free-living and symbiotic copepods. An overview of the basic functional morphology of the reproductive, locomotory and feeding systems will be presented, and the importance of the various sensory structures encountered in copepods will be discussed. Finally, a brief synopsis of the current ordinal classification system will be presented, including some recent developments in our phylogenetic understanding of the group.
This session will provide a brief overview of copepod morphology and diversity and introduce
participants to the impressive range of body forms and limb types across the Copepoda. Trainees will be introduced to the standardized terminology used in the description and identification of free-living and symbiotic copepods. An overview of the basic functional morphology of the reproductive, locomotory and feeding systems will be presented, and the importance of the various sensory structures encountered in copepods will be discussed. Finally, a brief synopsis of the current ordinal classification system will be presented, including some recent developments in our phylogenetic understanding of the group.
Lecture 1-2: Diversity of Copepod Life Cycles (Tutor: Rony Huys)
The ecological adaptability displayed by copepods is reflected in their tremendous morphological
plasticity which makes it difficult to formulate a rigorous diagnosis of the subclass Copepoda that is both informative yet sufficiently comprehensive to cover the bizarre parasites as well as the free-living forms. However, virtually all copepods have a stage in their life cycle, either the adult or one of the copepodid instars, exhibiting a cephalosome into which the maxilliped-bearing, first thoracic somite is incorporated and possessing at least two pairs of swimming legs, the members of which are linked by an intercoxal sclerite. The postembryonic development of copepods is divided into a naupliar phase
and a copepodid phase. Primitively, each phase comprises six stages but larval development and life cycles are highly variable and can be significantly abbreviated (0–6 nauplii; 1–6 copepodids). Most copepods hatch at the (ortho)nauplius stage, a simple larval form. Occasionally the first stage in the
life cycle is a metanauplius and some parasitic species are known to hatch as a copepodid. Typically, after six naupliar instars (often designated as NI, NII, NIII, etc.), the final nauplius stage moults into a copepodid stage, which resembles the adult but has a simple, unsegmented abdomen and only three
pairs of thoracic limbs (maxillipeds and legs 1–2). Copepodid stages are often referred to as CoI, CoII, etc. (or CI, CII, etc.). In some parasitic groups a stage following the infective copepodid (or one of the copepodid stages) is called the chalimus; it differs from the copepodid in its possession of a frontal filament that aids in attachment to the host. There are significant changes in body size, shape and appendages in the moult from NVI to CoI, collectively known as metamorphosis. Intermoult stages are important for tracing the origin and homology of larval structures between naupliar and copepodid
stages. Some parasitic copepods have an onychopodium or (transient) pupal stages in their life cycle. Participants will be introduced to the different larval types and the major differences between life cycles of free-living and symbiotic/parasitic copepods.
Lecture 1-3: Taxonomic Techniques for the Study of Copepods (Tutor: Rony Huys with assistance from other tutors)
This lecture aims at students becoming familiar with the basic methods used in taxonomic studies of copepods. The presentation will demonstrate how to produce adequate equipment to manipulate and
dissect copepods and how to prepare temporary and semi-permanent slides, focusing especially on the clearing, staining, mounting and dissecting techniques. The information content of taxonomic illustrations will be discussed, and detailed information on preparing line drawings, inking techniques and image editing will be provided.
Afternoon of Day 2 (27 May 2024)
Lecture 2-1: Morphology and Systematics of Marine Harpacticoida (Tutor: Rony Huys with assistance from other tutors)
Members of the order Harpacticoida are the benthic copepods par excellence and are best known for their ubiquity, diversity and hyperabundance in marine sediments, from the intertidal zone to the
deepest hadal trenches, spanning a vertical range in excess of 10,000 m. Harpacticoid copepods hold a similar position of pre-eminence in fresh and inland saline continental waters where they can be found in nearly every habitat and situation where sufficient moisture and organic matter are present.
Some species are no longer associated with the ancestral benthic environment and have exploited alternative habitats such as the open pelagic or the ephemeral habitat at the ice-water interface of polar and subpolar ice. Though relatively few species are known to be commensals, parasites or
otherwise associated, recent discoveries suggest that the true diversity of harpacticoids associated with invertebrate hosts has yet to be revealed. The taxonomy of harpacticoid copepods is in a constant state of flux. The order currently contains about 4,750 species in 700 genera and 53 families, hence it
would be impractical to cover all of them in detail. This lecture will provide an overview of harpacticoid copepod morphology and diversity, and introduce participants to the range of body forms and appendage types across the group. Preference will be given to the dominant families found in each of
the main habitats such as the interstitial and phytal environments, subtidal sandy sediments, mudflats and other silt-laden habitats, mangroves, the deep sea and the open pelagic. Morphological shortcuts based on characters that can be observed without dissection (e.g. antennule, maxilliped, leg 1) will be presented for the most important families but only relatively emphasis will be placed on families inhabiting freshwater. Finally, other non-harpacticoid taxa (e.g. Cyclopinidae, Cyclopidae, Canuellidae, Longipediidae, etc.) that frequently co-occur with harpacticoids in marine sediments will be briefly
presented.
Lecture 2-2: Ecology of Benthic Copepods in Extreme Environments (Tutor: Motohiro Shimanaga)
 Deepest hadal zones, hot hydrothermal vents, severe hypoxia in botom water, …—some environments can be defined as "extreme" if one or more of the physical or chemical variables are close to the limits that are tolerable by most life forms (cf. Zeppilli et al. 2018). If one extrapolates this definition, tidal ponds, which are easily accessible to humans, may also be perceived as an extreme environment for marine life due to their dramatic temporal changes in temperature and salinity.
Deepest hadal zones, hot hydrothermal vents, severe hypoxia in botom water, …—some environments can be defined as "extreme" if one or more of the physical or chemical variables are close to the limits that are tolerable by most life forms (cf. Zeppilli et al. 2018). If one extrapolates this definition, tidal ponds, which are easily accessible to humans, may also be perceived as an extreme environment for marine life due to their dramatic temporal changes in temperature and salinity.
Benthic Copepoda, primarily represented by harpacticoids, usually constitute the second-most abundant meiofaunal taxon after the nematodes. In general, they are more sensitive to environmental stresses than nematodes. Some specific copepod groups, however, have adapted to and can survive in such extreme environments. In this lecture, I will present the interesting ecological aspects associated with such copepod taxa, including their community structure, association with environmental factors, trophic ecology, distribution and genetic divergence, based on knowledge from recent studies, including our results.
Zeppilli, D., Leduc, D., Fontanier, C. et al. (2018) Characteristics of meiofauna in extreme marine
ecosystems: a review. Marine Biodiversity, 48, 35–71. https://doi.org/10.1007/s12526-017-0815-z
Lecture 3-1: Evolutionary Trends in Planktonic Calanoid Copepods (Tutor: Susumu Ohtsuka)
 The feeding strategies of calanoid copepods will be reviewed from morphological and evolutionary points of view. Calanoids can be classified into several types based on the structure of their feeding
appendages. The three different groups of particle-feeders differ primarily by the structure of their maxillae and maxillipeds, while carnivorous types are much more diversified in oceanic regions. In the oceanic genus Heterorhabdus and its related genera in the Heterorhabdidae, a venom-injection
system is employed to catch prey animals. Such system could have essentially evolved as a result of modifications of the feeding appendages and secretory glands of primitive particle-feeding heterorhabdids. The successive evolution of feeding strategies within the Heterorhabdidae will be demonstrated. Detritivorous Bradfordian families, such as the Scolecitrichidae and the Phaennidae, exhibit two types of chemosensory setae on the maxillae and maxillipeds, i.e. worm-like and brushlike setae, which may have different functions. TEM studies aimed at revealing the ultrastructure of these sensory setae showed that they are highly differentiated among the Bradfordian families. In addition to a variety of chemosensors, some deteritivorous copepods likely utilize different types of eyes to search for detrital matters containing bioluminescent bacteria. Complex combinations of the chemo- and optical sensors suggest niche separation in the Bradfordian families inhabiting oligotrophic oceanic waters.
The feeding strategies of calanoid copepods will be reviewed from morphological and evolutionary points of view. Calanoids can be classified into several types based on the structure of their feeding
appendages. The three different groups of particle-feeders differ primarily by the structure of their maxillae and maxillipeds, while carnivorous types are much more diversified in oceanic regions. In the oceanic genus Heterorhabdus and its related genera in the Heterorhabdidae, a venom-injection
system is employed to catch prey animals. Such system could have essentially evolved as a result of modifications of the feeding appendages and secretory glands of primitive particle-feeding heterorhabdids. The successive evolution of feeding strategies within the Heterorhabdidae will be demonstrated. Detritivorous Bradfordian families, such as the Scolecitrichidae and the Phaennidae, exhibit two types of chemosensory setae on the maxillae and maxillipeds, i.e. worm-like and brushlike setae, which may have different functions. TEM studies aimed at revealing the ultrastructure of these sensory setae showed that they are highly differentiated among the Bradfordian families. In addition to a variety of chemosensors, some deteritivorous copepods likely utilize different types of eyes to search for detrital matters containing bioluminescent bacteria. Complex combinations of the chemo- and optical sensors suggest niche separation in the Bradfordian families inhabiting oligotrophic oceanic waters.
Lecture 3-2: Taxonomy and Distribution of Planktonic Cyclopoid Copepods, especially Oithonidae (Tutor: Shuhei Nishida)
 Cyclopoid copepods of the family Oithonidae are among the most abundant, widely distributed, and ecologically important zooplankton. The species of this family are small, generally less than 1.5 mm
displaying varying levels of adaptability to physical and hydrological parameters. The role of Oithona was recognized in terms of their numerical contribution to the mesozooplankton and high turnover rates, proving to have a pivotal position in the food webs, microbial loops, and in carbon cycling. The Oithonidae, the family as a whole, is widely distributed in marine habitats, from estuaries to oceanic waters, from epipelagic to bathypelagic depths, and from tropical to polar regions. Currently four genera have been recognized in the family Oithonidae: Dioithona, Limnoithona, Oithona and Paroithona, of which Limnoithona is essentially limited to freshwater and, rarely, estuaries. However, still there is a debate on the status of Paroithona and Dioithona, necessitating further re-evaluation by molecular phylogenetic analysis based on an appropriate taxonomic coverage. In this lecture, the distribution and taxonomic issues in the family Oithonidae, and key characters for practical identification of their species will be presented.
Cyclopoid copepods of the family Oithonidae are among the most abundant, widely distributed, and ecologically important zooplankton. The species of this family are small, generally less than 1.5 mm
displaying varying levels of adaptability to physical and hydrological parameters. The role of Oithona was recognized in terms of their numerical contribution to the mesozooplankton and high turnover rates, proving to have a pivotal position in the food webs, microbial loops, and in carbon cycling. The Oithonidae, the family as a whole, is widely distributed in marine habitats, from estuaries to oceanic waters, from epipelagic to bathypelagic depths, and from tropical to polar regions. Currently four genera have been recognized in the family Oithonidae: Dioithona, Limnoithona, Oithona and Paroithona, of which Limnoithona is essentially limited to freshwater and, rarely, estuaries. However, still there is a debate on the status of Paroithona and Dioithona, necessitating further re-evaluation by molecular phylogenetic analysis based on an appropriate taxonomic coverage. In this lecture, the distribution and taxonomic issues in the family Oithonidae, and key characters for practical identification of their species will be presented.
Lecture 3-3: Taxonomy and Distribution of Planktonic Cyclopoid Copepods, especially Oncaeidae and Corycaeidae (Tutor: Hiroshi Ito)
 The families Corycaeidae and Oncaeidae both belong to the order Cyclopoida and can easily be recognized among marine zooplankton due to their unique morphology. Members of the family Corycaeidae are relatively large, have a pair of conspicuous lenses, and are typically found in the surface layer. An identification key that was constructed in the early 20th century is still in use today. On the other hand, many species of the family Oncaeidae have a body length of less than 0.5 mm and can be morphologically very similar. Their habitat ranges from the epipelagic zone to the deeper layers of the world’s oceans and from the equator to the polar regions. Since the 1960s the number of species has increased from about 20 to 100 and many unresolved taxonomical problems still remain. Corycaeids are typical carnivores that recognize and capture their prey by sight, while oncaeids are known to attach themselves to living Appendicularia, their abandoned houses and marine snow, and prey on them. Although data about their distribution, behavior, and physiological characteristics is continuously being accumulated, available information on the biology of both corycaeids and oncaeids is still significantly less than our current knowledge about members of the order Calanoida. It is expected that new research methods will reveal the unique role of these copepods in marine ecosystems. In this lecture, the distribution and taxonomic issues in the families Corycaeidae and Oncaeidae, and key characters for practical identification of their species will be presented.
The families Corycaeidae and Oncaeidae both belong to the order Cyclopoida and can easily be recognized among marine zooplankton due to their unique morphology. Members of the family Corycaeidae are relatively large, have a pair of conspicuous lenses, and are typically found in the surface layer. An identification key that was constructed in the early 20th century is still in use today. On the other hand, many species of the family Oncaeidae have a body length of less than 0.5 mm and can be morphologically very similar. Their habitat ranges from the epipelagic zone to the deeper layers of the world’s oceans and from the equator to the polar regions. Since the 1960s the number of species has increased from about 20 to 100 and many unresolved taxonomical problems still remain. Corycaeids are typical carnivores that recognize and capture their prey by sight, while oncaeids are known to attach themselves to living Appendicularia, their abandoned houses and marine snow, and prey on them. Although data about their distribution, behavior, and physiological characteristics is continuously being accumulated, available information on the biology of both corycaeids and oncaeids is still significantly less than our current knowledge about members of the order Calanoida. It is expected that new research methods will reveal the unique role of these copepods in marine ecosystems. In this lecture, the distribution and taxonomic issues in the families Corycaeidae and Oncaeidae, and key characters for practical identification of their species will be presented.
Lecture 3-4: Distribution of Planktonic Calanoid Copepods (Tutor: Atsushi Yamaguchi)
 As habitats of planktonic calanoid copepods, neritic and oceanic are classified as the horizontal category. Regarding latitudinal direction, north to south or high latitude to low latitude are also
important. From a vertical point of view, the near-surface neuston layer, epipelagic, mesopelagic, bathypelagic, and abyssopelagic layers can be recognized. Horizontal distribution of planktonic calanoid copepods is mainly governed by their life cycle patterns. In neritic areas, calanoid communities are typically dominated by species that produce resting eggs which survive in muddy sediments. In oceanic areas, the dominance of large-sized copepods, which have resting or diapause phases in the deeper layers, is also common. In terms of latitudinal changes, the higher latitude seas of both the Northern and Southern Hemispheres are dominated by large-sized species which display resting phases in deep layers.
As habitats of planktonic calanoid copepods, neritic and oceanic are classified as the horizontal category. Regarding latitudinal direction, north to south or high latitude to low latitude are also
important. From a vertical point of view, the near-surface neuston layer, epipelagic, mesopelagic, bathypelagic, and abyssopelagic layers can be recognized. Horizontal distribution of planktonic calanoid copepods is mainly governed by their life cycle patterns. In neritic areas, calanoid communities are typically dominated by species that produce resting eggs which survive in muddy sediments. In oceanic areas, the dominance of large-sized copepods, which have resting or diapause phases in the deeper layers, is also common. In terms of latitudinal changes, the higher latitude seas of both the Northern and Southern Hemispheres are dominated by large-sized species which display resting phases in deep layers.
On the other hand, calanoid communities in lower latitude seas are dominated by small-sized copepods, show a high species diversity and their feeding modes vary according to depth. In the epipelagic layers copepod communities are typically dominated by the particle feeders while those of the mesopelagic layers are predominantly represented by detritivores and carnivores. The species diversity of the planktonic copepods is known to be highest in the mesopelagic layers of various locations in the world’s oceans which may be a reflection of their divergent feeding modes.
Morning of Day 3 (28 May 2024)
Lecture 4-1: Symbiotic Copepods Using Invertebrate Hosts: Diversity and Adaptation (Tutor: Rony Huys)
Copepods utilise a spectacular variety of invertebrate metazoans as hosts, from sponges to urochordates. Relatively few species of copepods make use of host groups such as the Nemertea,
Platyhelminthes, Bryozoa, Phoronida, Brachiopoda, Hemichordata and Sipuncula. However, they are very commonly associated with sponges, the entire range of cnidarian groups from the hard corals and sea anemones to the medusae and siphonophores, most echinoderm and molluscan groups, both solitary and colonial tunicates, and to a lesser extent, crustaceans and annelids (including the former phyla Echiura and Vestimentifera). The main families utilising invertebrate host phyla will be introduced and their usage of host microhabitats reviewed. The body form of copepods parasitic on marine invertebrates is astonishingly varied. Many retain the basic cyclopiform body but some of the most extreme examples of secondary reduction in body segmentation combined with the loss of paired limbs are found in copepods from invertebrate hosts. A surprising variety of species also possess a system of rootlets which penetrate host tissues. Students will be introduced to where to find symbiotic copepods, their adaptations to the parasitic mode of life, and the characters that are
most useful in their identification. Key aspects of the biology of a selection of the most important taxa and methods of collecting and extracting copepods from their invertebrate hosts will also be summarised.
Lecture 4-2: Parasitic Copepods Found on Egg-masses of Lobsters (Tutor: Kaori Wakabayashi)
 Copepods in the family Nicothoidae exclusively parasitize crustacean hosts and many of them are egg mimics. They typically associate with the brood pouch of peracaridans or external egg masses of decapods and resemble the eggs of their hosts in size and in their globular body form. Members of the genus Choniomyzon are known to associate with the external egg mass of decapods: three species have been described from achelate lobsters and another species from a crab belonging to the genus Libinia. The reproductive strategy appears to be different between the lobster egg symbionts and the species associated with the crab host. The adult female of the former three species carries several egg sacs, each containing 10–20 embryos. Nauplii hatch from the egg sacs while these are still carried by the maternal female. On the other hand, the adult female of the crab-associated species produces several egg sacs, each containing only a single embryo, which are deposited onto the host's eggs until the nauplii hatch. Copepodid I was observed on the body of the decapod host, either on its surface or on the gills, but never on the egg masses. This suggests that infection takes place at copepodid I stage after dispersal during the naupliar phase and that moulting into copepodid II is delayed until the host becomes ovigerous. In this lecture, the current understanding of the taxonomy and the life cycle of Choniomyzon is summarized and the methods employed in finding and observing each developmental stage of the copepods are explained.
Copepods in the family Nicothoidae exclusively parasitize crustacean hosts and many of them are egg mimics. They typically associate with the brood pouch of peracaridans or external egg masses of decapods and resemble the eggs of their hosts in size and in their globular body form. Members of the genus Choniomyzon are known to associate with the external egg mass of decapods: three species have been described from achelate lobsters and another species from a crab belonging to the genus Libinia. The reproductive strategy appears to be different between the lobster egg symbionts and the species associated with the crab host. The adult female of the former three species carries several egg sacs, each containing 10–20 embryos. Nauplii hatch from the egg sacs while these are still carried by the maternal female. On the other hand, the adult female of the crab-associated species produces several egg sacs, each containing only a single embryo, which are deposited onto the host's eggs until the nauplii hatch. Copepodid I was observed on the body of the decapod host, either on its surface or on the gills, but never on the egg masses. This suggests that infection takes place at copepodid I stage after dispersal during the naupliar phase and that moulting into copepodid II is delayed until the host becomes ovigerous. In this lecture, the current understanding of the taxonomy and the life cycle of Choniomyzon is summarized and the methods employed in finding and observing each developmental stage of the copepods are explained.
Lecture 4-3: An Overview of Parasitic Copepods on Vertebrate Hosts (Tutor: Danny Tang)
 Copepods are common parasites of marine and freshwater fishes and are known to cause diseases in finfish aquaculture. They may also serve as useful bioindicators of host dispersal, host phylogeny and
host population structure. Participants will be introduced to the morphological diversity, host microhabitats, modes of attachment and life histories of representative piscicolous copepods. Emphasis will be placed on the Ergasilidae, Chondracanthidae, and Taeniacanthidae for the poecilostome cyclopoids and on the Caligidae, Pennellidae and Lernaeopodidae for the Siphonostomatoida.
Copepods are common parasites of marine and freshwater fishes and are known to cause diseases in finfish aquaculture. They may also serve as useful bioindicators of host dispersal, host phylogeny and
host population structure. Participants will be introduced to the morphological diversity, host microhabitats, modes of attachment and life histories of representative piscicolous copepods. Emphasis will be placed on the Ergasilidae, Chondracanthidae, and Taeniacanthidae for the poecilostome cyclopoids and on the Caligidae, Pennellidae and Lernaeopodidae for the Siphonostomatoida.
Lecture 4-4: How do Caligids Adhere to Fish? (Tutor: Susumu Ohtsuka)
Caligus adheres to the body surface of host fish with a cephalothoracic sucker, using legs 2 and 3 in well-organized manners. Lunules specific Caligus and some related genera have a vital function in the initial step of adhering and contain a highly unique endocuticule ultrastructure that may behave like active matter and serve as the actuating mechanism. Cuticular membranes bordering the cephalothorax have a unique endocuticule with an undulating dorsal surface and a smooth ventral surface. A high-speed camera revealed that this undulation likely facilitates rapid automatic application of the sucker to the substrate. The cuticular membranes on the posterior margin of the first exopodal segment of leg 2 have a specialized endocuticle with tubules each surrounded by fine fibers. This reinforcement helps them to generate a posteriorly-directed jet of water. Opening-closing of these membranes is controlled by postero-anterior motion of the distal exopodal segments of leg 2. The outer cuticular membrane of leg 3 is simple, presumably effected by powerful extrinsic muscles. The consistency of sucker morphology within Caligus implies a highly stereotyped attachment behavior on fish skin covered with mucous. In short, Caligus can convert simple antero-posteior movements of legs into closure of the sucker.
[based on: Ohtsuka et al., (2021), The cephalothoracic sucker of sea lice (Crustacea: Copepoda: Caligidae): The functional importance of cuticular membrane ultrastructure, Arthropod Structure & Development, 62, 101046.]
Lecture 4-5: Taxonomy of Pennellidae and Chondracanthidae in Asian Waters (Tutor: Panakool Thambun Aneesh)
 Globally, information on parasitic crustaceans is patchy, and taxonomic knowledge of marine fish parasitic copepods in Asian waters has not progressed significantly since 1985. Taxonomic knowledge
of copepods infesting commercially important fish has been comparatively well documented during the last few decades, however, only a limited number of taxonomic publications are available on noncommercial and deep-sea fishes. During this lecture, a brief review will be presented on the morphology and diversity of two highly transformed fish parasitic copepod families, the Pennellidae Burmeister, 1835 and the Chondracanthidae Milne Edwards, 1840, with special reference to taxa occurring in Indian waters. The mesoparasitic copepod family Pennellidae currently accommodates 145 species in 24 valid genera. The general structure of appendages and different body forms within this family will be discussed. The Chondracanthidae is one of the most speciose copepod families and accommodates around 190 species in 50 valid genera, all of which exclusively utilize fishes as hosts. Among them, 27 genera are monotypic and their validity must be assessed with molecular sequence data. Only two genera include more than ten species, Acanthochondria Oakley, 1930 with 54 valid species, and Chondracanthus Delaroche, 1811 with 41 valid species. Typically, adult females show a higher modification of the body and its appendages than their corresponding males which often have a reduced body size (dwarf/pygmy males). The general structure of appendages and different body forms within the family Chondracanthidae will be discussed and their evolutionary trends reviewed.
Globally, information on parasitic crustaceans is patchy, and taxonomic knowledge of marine fish parasitic copepods in Asian waters has not progressed significantly since 1985. Taxonomic knowledge
of copepods infesting commercially important fish has been comparatively well documented during the last few decades, however, only a limited number of taxonomic publications are available on noncommercial and deep-sea fishes. During this lecture, a brief review will be presented on the morphology and diversity of two highly transformed fish parasitic copepod families, the Pennellidae Burmeister, 1835 and the Chondracanthidae Milne Edwards, 1840, with special reference to taxa occurring in Indian waters. The mesoparasitic copepod family Pennellidae currently accommodates 145 species in 24 valid genera. The general structure of appendages and different body forms within this family will be discussed. The Chondracanthidae is one of the most speciose copepod families and accommodates around 190 species in 50 valid genera, all of which exclusively utilize fishes as hosts. Among them, 27 genera are monotypic and their validity must be assessed with molecular sequence data. Only two genera include more than ten species, Acanthochondria Oakley, 1930 with 54 valid species, and Chondracanthus Delaroche, 1811 with 41 valid species. Typically, adult females show a higher modification of the body and its appendages than their corresponding males which often have a reduced body size (dwarf/pygmy males). The general structure of appendages and different body forms within the family Chondracanthidae will be discussed and their evolutionary trends reviewed.
Afternoon of Day 5 (30 May 2024)
Lecture 5-1: Copepods in Fresh Water: Systematics and Morphology of Continental Copepods, a Quick Overview through all Continental Habitats that Copepods can Occupy (Tutor: Nancy F. Mercado-Salas)
 Freshwater habitats house an extraordinary diversity in comparison with the small proportion of the Earth‘s surface they occupy. This great diversity results from the variability in environmental
characteristics of freshwater habitats, each with their own set of adapted populations. In the case of freshwater environments, the larger and more permanent water bodies have received most attention by researchers and small, temporary and peripheral aquatic habitats have been largely ignored in
terms of biodiversity surveys. In comparison with marine environments, relatively few families of copepods are present in continental ecosystems; however, in terms of diversity the most speciose copepod families are found in such environments (Cyclopidae, Diaptomidae, Canthocamptidae).
Freshwater habitats house an extraordinary diversity in comparison with the small proportion of the Earth‘s surface they occupy. This great diversity results from the variability in environmental
characteristics of freshwater habitats, each with their own set of adapted populations. In the case of freshwater environments, the larger and more permanent water bodies have received most attention by researchers and small, temporary and peripheral aquatic habitats have been largely ignored in
terms of biodiversity surveys. In comparison with marine environments, relatively few families of copepods are present in continental ecosystems; however, in terms of diversity the most speciose copepod families are found in such environments (Cyclopidae, Diaptomidae, Canthocamptidae).
Participants will be introduced to the major continental families and the morphological characters used to distinguish them. The main groups inhabiting continental environments will be briefly presented, including brackish and coastal systems (Euryteidae, Halicylopinae), freshwater plankton (Cyclopidae, Diaptomidae, Canthocamptidae), subterranean and anchialine ecosystems (Cyclopidae, Halicyclopinae, Parastenocarididae, Chappuisiidae, Speleophriidae, Pseudocyclopidae), and finally, semi-terrestrial and other cryptic habitats (Cyclopidae, Canthocamptidae, Phyllognathopodidae). A discussion of the phylogenetic position of continental taxa, including morphology and molecular work, will be presented.
Lecture 5-2: Taxonomy and Phylogeny of Freshwater Cyclopoida in Asia (Tutor: Ko Tomikawa)
 The Cyclopidae is a cosmopolitan family of Copepoda characterized by a high species diversity in a variety of freshwater environments, including surface water and groundwater. So far, over 800 species in 70 genera have been described worldwide, of which 72 species in 19 genera occur in Japan. Members of the Cyclopidae perform an important role in the food chain of freshwater ecosystems since they act either as primary consumers of algae and/or as secondary consumers of Cladocera. Recently, there has also been concern about the impact of some species of this family on the ecosystems as introduced species. In this lecture, I will introduce participants to the species diversity and identification of major groups of Cyclopidae in Japan and other Asian countries.
The Cyclopidae is a cosmopolitan family of Copepoda characterized by a high species diversity in a variety of freshwater environments, including surface water and groundwater. So far, over 800 species in 70 genera have been described worldwide, of which 72 species in 19 genera occur in Japan. Members of the Cyclopidae perform an important role in the food chain of freshwater ecosystems since they act either as primary consumers of algae and/or as secondary consumers of Cladocera. Recently, there has also been concern about the impact of some species of this family on the ecosystems as introduced species. In this lecture, I will introduce participants to the species diversity and identification of major groups of Cyclopidae in Japan and other Asian countries.
Lecture 6: From Phenotypes to Genotypes and Back: Towards an Integrated Evaluation of Biodiversity in Calanoid Copepods (Tutor: Iole Di Capua)
 Copepods, the most abundant metazoans on Earth, exert significant influence over marine planktonic ecosystems in both abundance and diversity. Since the late 19th century, over 2,600 marine calanoid copepod species have been identified across various basins. In recent years, molecular genetic techniques have gained prominence for detecting, discriminating, and identifying these copepod species, offering a valuable complement to traditional morphological methods.
Copepods, the most abundant metazoans on Earth, exert significant influence over marine planktonic ecosystems in both abundance and diversity. Since the late 19th century, over 2,600 marine calanoid copepod species have been identified across various basins. In recent years, molecular genetic techniques have gained prominence for detecting, discriminating, and identifying these copepod species, offering a valuable complement to traditional morphological methods.
Increasingly, molecular approaches, particularly DNA barcoding, have played a pivotal role in refining our understanding of copepod diversity. Utilizing highly conserved regions of the mitochondrial cytochrome c oxidase I (COI) gene, DNA barcoding has proven to be a precise and reliable tool for identification. Although calanoid copepods stand as one of the extensively studied groups, using molecular approaches only 104 out of the 2,600 identified species (4%) presently have COI barcodes, primarily from the order Calanoida (42%).
This lecture aims at highlighting the efficacy of DNA barcodes in the identification of known, newly discovered, and cryptic calanoid species. It also seeks to elucidate the current state of knowledge regarding calanoid taxonomic diversity by integrating genotypic and phenotypic approaches.
Lecture 7: Copepods as Hosts for other Crustaceans: an Overview (Tutor: Alexandra Savchenko)
 Nearly half of the known copepod species have entered into symbiotic relationships with a wide range of metazoan hosts. Copepods themselves are known to serve as hosts for a number of unicellular and
multicellular parasites, including other crustaceans. During the workshop participants will be introduced to two groups of marine parasitic crustaceans which utilize copepods either as intermediate (Epicaridea) or definitive hosts (Tantulocarida). Epicaridean isopods (families Bopyridae, Cryptoniscidae, Dajidae, Entoniscidae) infect marine planktonic copepods at the microniscus larval stage. This larva has particular morphological adaptations to attach itself externally to a very motile host, where it moults up to six times to become the next infective stage – the cryptoniscus larva – which leaves the copepod to seek the definitive host. In all known epicaridean life cycles the definitive hosts are free-living or parasitic crustaceans, including decapods, euphausiids, mysids, barnacles (including Rhizocephala), ostracods and other isopods.
Nearly half of the known copepod species have entered into symbiotic relationships with a wide range of metazoan hosts. Copepods themselves are known to serve as hosts for a number of unicellular and
multicellular parasites, including other crustaceans. During the workshop participants will be introduced to two groups of marine parasitic crustaceans which utilize copepods either as intermediate (Epicaridea) or definitive hosts (Tantulocarida). Epicaridean isopods (families Bopyridae, Cryptoniscidae, Dajidae, Entoniscidae) infect marine planktonic copepods at the microniscus larval stage. This larva has particular morphological adaptations to attach itself externally to a very motile host, where it moults up to six times to become the next infective stage – the cryptoniscus larva – which leaves the copepod to seek the definitive host. In all known epicaridean life cycles the definitive hosts are free-living or parasitic crustaceans, including decapods, euphausiids, mysids, barnacles (including Rhizocephala), ostracods and other isopods.
The Tantulocarida is a distinct class of minute parasitic microcrustaceans that exclusively utilize other crustaceans as hosts. Crustacean host categories include Amphipoda, Isopoda, Ostracoda, Cumacea and Tanaidacea, however, most commonly tantulocaridans are found on (hyper)benthic harpacticoid and other copepods. Tantulocaridans display a complex dual life cycle. The infective stage known as the tantulus larva locates a suitable host and becomes permanently attached to it by means of a special proteinous glue. The tantulus feeds using a special absorptive rootlet system which grows inside the host through a tiny opening made by the unpaired cephalic stylet. Further metamorphosis which proceeds without typical moulting leads to the release of the next generation of tantuli (parthenogenesis) or free swimming non-parasitic sexual stages.
Finally, the symbiotic association of pedunculate barnacles (Conchoderma spp.) with pandarid and pennellid parasitic copepods (Siphonostomatoida) on marine fish and mammal hosts will be briefly discussed.
Evening of Day 5 (30 May 2024)
Lecture 8: An Introduction to the Principles of Zoological Nomenclature (Tutor: Rony Huys)
The International Code of Zoological Nomenclature (ICZN or The Code) is a widely accepted convention in zoology that rules the formal scientific naming of organisms treated as animals. The rules principally regulate (1) How names are correctly established in the frame of binominal nomenclature, (2) Which name must be used in case of name conflicts, and (3) How scientific literature must cite names. The rules and recommendations have one fundamental aim: to provide maximum universality and continuity in the naming of all animals, except where taxonomic judgement dictates otherwise. The Code is meant to guide only the nomenclature of animals, while leaving zoologists
freedom in classifying new taxa. Students will be introduced to the basic principles of zoological nomenclature including availability, validity, priority, synonymy and homonymy of taxonomic names.
Training Cruise
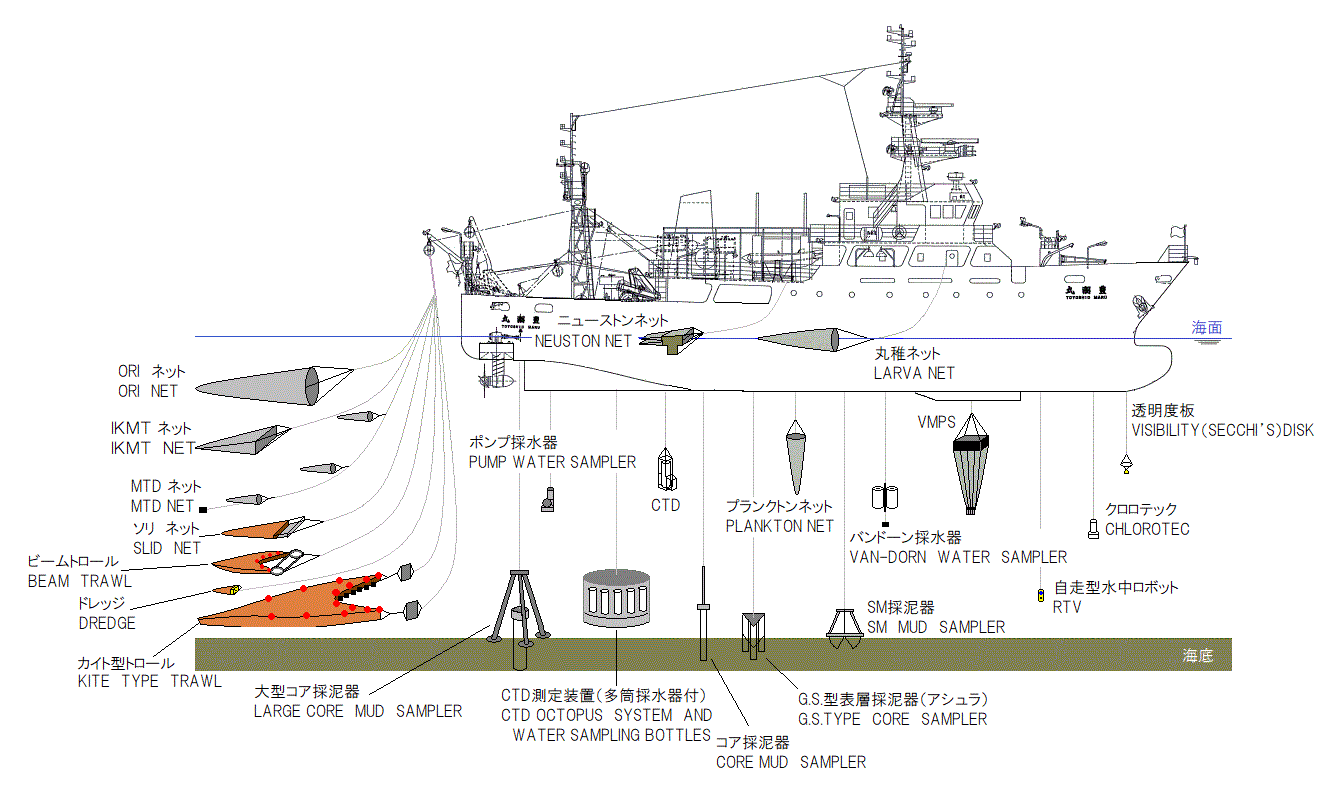
During the Pre-conference Workshop, all trainees and tutors will have the opportunity to learn how to operate a variety of sampling gears on board the TRV Toyoshio-Maru: CTD, Secci disc, plankton net, neuston net, sledge net, fish larva net, Smith-McIntyre grab, core sampler, fishing rod etc. These gears will be deployed at some stations in the Seto Inland Sea, Japan. In addition, participants will be able to experience 24 h life on board the vessel.
Schedule
Group A: 28 and 29 May 2024
Group B: 29 and 30 May 2024
Practical Session
 Trainees will learn and experience the skills of fixation, dissection, and observation of copepod specimens. Individual tutors will demonstrate how to examine and identify specimens depending on the target group. The practical sessions will provide guided dissections of selected tutors’ material (free-living and parasitic examples). Trainees are encouraged to bring their own copepod specimens if they wish to identify them for their personal studies. In one’s own interest it is strongly advised that successful applicants contact their most appropriate tutor(s) to discuss problems beforehand so that relevant documentation and literature can be provided. Given the vast amount of literature on copepods do not expect tutors to have access to every single species description during the workshop. Prior consultation with the tutors is therefore recommended if not essential. A variety of planktonic, benthic, and symbiotic copepods collected during the training cruise will also be available for study. Each trainee will receive a free copy of An Introduction to Copepod Diversity by Boxshall & Halsey (2004) which will facilitate the identification of copepods to family or genus level. Biological, stereo, and scanning electron microscopes will be available in the laboratory. It is highly recommended that participants bring their own dissection tools. Trainees may also wish to discuss molecular techniques required to carry out phylogenetic analyses of copepods.
Trainees will learn and experience the skills of fixation, dissection, and observation of copepod specimens. Individual tutors will demonstrate how to examine and identify specimens depending on the target group. The practical sessions will provide guided dissections of selected tutors’ material (free-living and parasitic examples). Trainees are encouraged to bring their own copepod specimens if they wish to identify them for their personal studies. In one’s own interest it is strongly advised that successful applicants contact their most appropriate tutor(s) to discuss problems beforehand so that relevant documentation and literature can be provided. Given the vast amount of literature on copepods do not expect tutors to have access to every single species description during the workshop. Prior consultation with the tutors is therefore recommended if not essential. A variety of planktonic, benthic, and symbiotic copepods collected during the training cruise will also be available for study. Each trainee will receive a free copy of An Introduction to Copepod Diversity by Boxshall & Halsey (2004) which will facilitate the identification of copepods to family or genus level. Biological, stereo, and scanning electron microscopes will be available in the laboratory. It is highly recommended that participants bring their own dissection tools. Trainees may also wish to discuss molecular techniques required to carry out phylogenetic analyses of copepods.
Hatchery Tour
On the last day (31 May 2024) participants will visit the Hiroshima Prefectural Farming Fisheries Center located near the venue, where we will observe farming of seeds of some commercially important fish and invertebrates such as red sea bream, swimming crab and oyster.
Hiroshima Prefectural Farming Fisheries Center



